![]()
AUTOR: Milan Radulović (milan.radulovic@poreklo.rs)
Objavljeno: 24.8.2019.
PDF
Opšte napomene:
Haplogrupa L nastala je iz predačke haploskupine LT, kada je proistekla iz nje prije 42.000 godina, dok je zajednički predak svih L grana živio prije 24.000 godina na prostoru Tadžikistana. Dijeli se na L1 i L2 haplogrupu. Haplogrupa L1 grana se na tri velike grane, L1a1-M27 koja je karakteristečna za prostor južne i jugoistočne Azije, L1a2-M357 karakterističnu za centralnu i zapadnu Aziju i L1b koja ima svoja žarišta u Evropi, Maloj Aziji i Levantu. L2 predstavlja jednu od najređih haplogrupa na svijetu. Sreće se mahom u Evropi, a registrovana je još i u jugoistočnoj Anadoliji, Kakvakzu, Rusiji (Tatarstanu). Haplogrupa L može se povezati sa Civilizacijom doline Inda, Kura-Arakses i Majkop kulturom; Dravidima, Paštunima, Beludžima i drugim južno-azijskim narodima. Takođe se može povezati i sa Grcima Ponta i Anadolije, posebno L1b linije.
Porijeklo i migracije
Haplogrupa L nastala je prije 42.600 godina, a živjela je na ivici izumiranja do prije 24.100 godina kada je živio zajednički predak svih L linija (YFull, 2019). Za mjesto porijekla smatra se Tadžikistan, odnosno planinski vijenac Pamir, navode Mahal i Matsoukas (Mahal & Matsoukas, 2017). Dalje se grana na L1 i L2 haplogrupu. Grana L1a razvila se na području između zapadnog Pakistana i Indije, dok su L1b i L2 rano migrirali u pravcu zapadne Azije i Zakavkazja.
Neolit i eneolit
L1a-M2481 tokom neolita i eneolita
Haplogrupa L1 grana se na dvije grane L1a i L1b, koje su formirane prije 18.000 godina. Tokom neolita i eneolita grane L1a, M357 i M27 zastupljene su na širokom prostoru od zapadnog Irana i Zakavkazja do južne Indije, sa primarnim žarištem u zapadnom Pakistanu i Indiji. Sa širenjem poljoprivrede iz pravca centralnog Zagorsa, postaju prvi poljoprivrednici južne Azije. Predstavljajući osnovnu haplogrupu visoko razvjene Mergar kulture (7.000-3.300. p.n.e.). Najveću raznovrsnost haplotipova bilježi u Indiji gdje je prema Trivediju i srardnicima (Trivedi, Sahoo, Singh, Bindu, & Banerjee, 2007) raznovrsnost ±0.998. Što zajedno sa visokom frekvencijom ide u prilog starosti haplogrupe na području južne Azije. Tokom eneolita proširili su se do Zakavkazja gdje je nađena L1a1-M27 u tri uzorka u Areni pećini u Jermeniji, I1634 (4330-4060 p.n.e.), I1632 (4230-4000 p.n.e.), I1407 (4350-3500 p.n.e.), datovanih u eneolit (Lazaridis, et al., 2016).
Sa nastankom Civilizacije doline Inda 3.300. p.n.e. stanovništvo se iz Mergara seli ka dolini Inda zasnivajući velike gradove Mohendžo-daro, Harapa i Lotal. Gdje je L1a bila pretežna haplogrupa, pored J2-M172.
L1b-M317 tokom neolita i eneolita
Haplogrupa L1b koju definiše snip M317 takođe je kao i L1a nastala na prostoru zapadnog Pakistana u dolini Inda, gdje je i danas prisutna sa oko 1,14% (Sengupta, et al., 2006). No za razliku od L1a linija, sreće se uglavnom u društvenim skupinama koje govore indo-evropske jezike, zaključujući prema rezultatima iz pomenutog rada i rezultatima iz studije Di Kristofara (Di Cristofaro, et al., 2013). Zajednički predak svih L1b muškaraca živio je prije 12.700 godina. Primarni centar haplogrupe je na Levantu gdje je najveća zastupljenost i raznolikost, dok je značajan dio populacije danas prisutan u Evropi, što nas dovodi do zaključka da je L1b-M317 imala rano migracijsko razdvajanje od L1a linija. Zaključujući prema današnjoj rasprostranjenosti, učestalost i raznolikosti L1b haplogrupe, razvila se na području sjevernog Iraka.
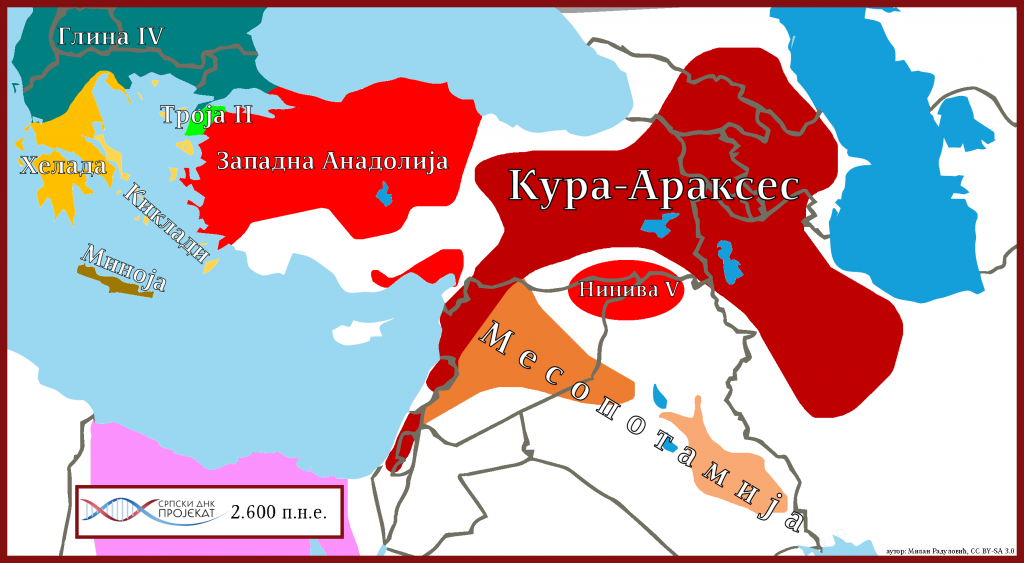
Kako za sada nije nađena u drevnoj DNK prvih farmera Levanta, te kako izostaje iz neolitskih uzorka Anadolije i kako je imala prilično rano razdvajanje od drugih L linija, nameće se pitanje vremenskog okvira širenja populacije. Pretpostavljajući sa druge strane da je L1b-M317 bila jedna od linija Kura-Arakses (3.400-2.000 p.n.e.) kulture (Hay, 2018) i uzmajući u obzir da su bratske L1a i L2 linije nađene u uzorcima iz eneolitskog Zakavkazja (4.300 p.n.e.) i kasnog Majkopa (3.400 p.n.e.), možemo zaključiti da su L1b muškarci započeli demografsko širenje krajem eneolita na području između Zakavkazja i sjevera Iraka.
L2-L595 tokom neolita i eneolita
Predstavlja jednu od najređih haplogrupa na svijetu. Za sada je pronađena kod nekolicine testiranih. Proistekla je iz očinske L-M20 skupine prije 23.000 godina, na području Tadžikistana. Rano je migrirala prema Kavkazu nezavisno od L1 linija gdje se razvijala i formirala primarno žarište. U starim kostima pronađena je na lokalitetu Tepe Hisar na sjeveroistoku Irana, lokalitetu koji predstavlja kasno eneolitsko i rano bronzanodobsko nalazište.
Bronzano i gvozdeno doba
Tokom bronzanog doba dolazi do značajnijeg demografskog širenja L-M20 grana prema centralnoj i zapadnoj Aziji. Značajan uticaj na ekspanziju haplogrupe L-M20 imale su sledeće kulture: Baktrijsko-Margianski arheološki kompleks sa područja istočnog Turkmenistana, južnog Uzbekistana, sjevernog Afganistana i zapadnog Tadžikistana; Civilizacija doline Inda iz Pakistana; te Kura Arakses i Majkop sa prostora Zakavkazja.
L1a-M2481 tokom bronzanog i gvozdenog doba
Epicentar M2481 tokom bronzanog doba bio je na prostoru Baktrijsko-Margianskog arheološkog kompleksa (BMAK) koji se kako smo već ranije pomenuli nalazio na prostoru između istočnog Turkmenistana, južnog Uzbekistana, sjevernog Afganistana i zapadnog Tadžikistana. Pored današnje visoke zastupljenosti haplogrupe na pomenutom području i njene raznovrsnosti (raznovrsnosti haplotipova). Na žarišno prisustvo tokom bronzanog doba upućuju i nalazi starih kostiju odnosno drevne DNK.
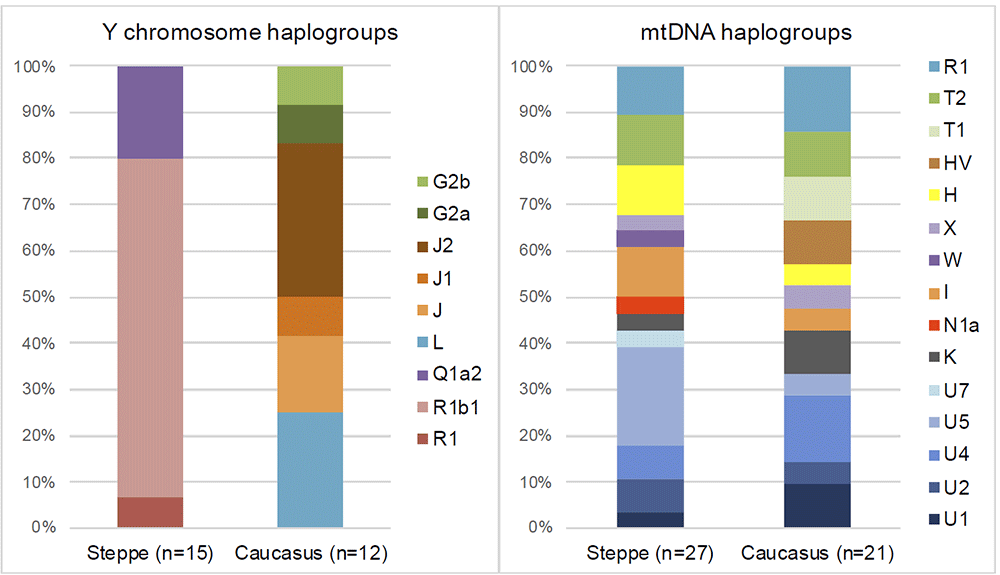
Prema nalazima iz studije Narasimhana i saradnika iz 2018. godine (Narasimhan, et al., 2018), u testiranoj muškoj populaciji BMAK, od 18 skeleta, 2 su L1a haplogrupe. Uzorak I5604 datiran je u period 1880-1697 p.n.e., a I4285 u 1873-1661 p.n.e.. U populaciji BMAK nađena su 4 uzorka J2a, po dva uzorka E1b1, R2a, G i J haplogrupe i po jedan R1b1, J1, R, T. Iz studije se zaključuje da je evidentno kretanje populacije u smijeru zapad-istok odnosno iz pravca Anadolije i Levanta ka istoku. Nadalje, zaključuje se da je populacija BMAK većinski lokalnog eneolitskog kontinuiteta i da nema bitnijeg stepskog uticaja. Još se navodi, da grupacije koje okružuju BMAK nemaju stepsko porijeklo do 2.100 p.n.e., te da se uticaj stepskih populacija evidentira tek u periodu imeđu 2.100-1.700 p.n.e..
Tako da nalazi koje dobijamo iz „drevne DNK“ sugerišu da su nosioci L-M20 haplogrupe započeli širenje u centralnu Aziju tek u periodu posle 1.700. p.n.e., i da se nisu nalazili u korpusu stepske populacije eneolita.
Dok su se kretanja L1a-M2481 ka unutrašnjosti Indije dešavala tokom mezolita smatra Saho (Sahoo, et al., 2006). No kako je L-M20 prisutan i u indo-evropskim kastama do oko 7%, jedan dio L1a svakako je stigao sa indo-evropskim migracijama.
![]()
Što se tiče Zakavkazja, osvrćući se na eneolitske uzorke iz Jermenije, M2481>M27, te L2 iz Majkopa, vrlo je moguće da su L-M20 (M2481, L595) muškarci bili jedni od nosilaca majkopske kulture i da su se u kasnijoj složenoj interakciji sa Kura-Arakses kulturom širili Anadolijom. Tako da su krajem bronzanog ili početkom gvozednog doba prešli u Evropu.
L1b-M317 tokom bronzanog i gvozdenog doba
Kada je riječ o L1b linijama pored klasičnih metoda, oslonjenih na istoriju, zastupljenost i raznovrsnost haplogrupe, mi još uvijek ne raspolažemo nalazima drevne DNK što u izvjesnoj mjeri otežava kvalitetan pristup razumjevanju migracionih odnosa. Stoga smo pri donošenju zaključaka pored ostalog značajnije oslonjeni na opšti odnos i migracije haplogrupa sa kojima su L1b-M317 u dodiru.
Razvila se prije 12 hiljada godina na podrčju između Zakavkazja i Mesopotamije, na šta upućuje najveća raznovrsnost i zastupljenost haplotipova, o čemu je bilo riječi.
Tokom bronzanog doba učestvovali su u populaciji Kura-Arakses (3.400-2.000 p.n.e.) kulture, koja se protezala od istočne Anadolije i Plodnog polumjeseca na istoku do Irana na zapadu. Pripadnici ove kulture bili su pretežno J2a-L26, J1, G2a-L293, R1b, J2b, L1b, T1a-P77 Y-DNK haplogrupa (Radulović i Novaković, 2018).
![]()
Kako je Kura-Arakses identifikovana kao kultura od posebnog značaja za L1b haplogrupu, shodno tome važno je napomenuti da je ova kultura bila polu ratničkog ili relativno miroljubivog tipa i kao takva u literaturi je označena. Ona je imala uticaja na kulture Anadolije, te na ostrvo Kipar navode Alizadeh i saradnici (Alizadeh, Eghbal, & Samei, 2015), a još je moguće imala i uticaja na Krit preko Anadolije. Prema brojnim autorima upravo je Kura-Arakses uticala na kolaps Uruka (4.000-3.100 p.n.e.) (Kohl, 2007), što je moguće uticalo na širenje L1b populacije u pravcu jugoistoka, ali i Arabijskog poluostrva.
![]()
Tako da odgovore na migracione procese L1b populacije ka Evropi i mediteranskom pojasu treba tražiti prevashodno u ekspanziji Kura Arakses kulture. Još je bitno imati u vidu da su na prostoru Kavkaza, odnosno na prostoru Majkop kulture pronađeni pripadnici L1a i L2 haplogrupa (Lazaridis, et al., 2016; Wang, et al., 2018; Rajevac, 2019), a znajući da je Kura Arakses geografski u dodiru sa prostorom na kome se nalazila Majkop kultura (3.700-3.300 p.n.e.); sasvim je za pretpostaviti da su i L1b muškarci bili prisutni u Majkop kulturi, a čiji su predstavnici pretežno bili J2a-L26, J1, L1a, G2, L2, J2b muškarci.
L2-L595 tokom bronzanog i gvozdenog doba
Kretanje L595 tokom istorije nešto je teže ispratiti usled malobrojnosti skupine. Naime, za sada je testirano svega 15-ak ljudi koji su pripadnici ove rijetke haplogrupe. Za potporu teza koje ćemo iznijeti oslonićemo se na nalaze drevne DNK, odnose haplogrupa sa kojima su u dodiru i na trenutnu rasprostranjenost.
U radu Vanga i saradnika indentifikovani su tri uzorka L haplogrupe koji se vezuju za kasni Majkop, a za koje Rajevac (Rajevac, 2019) precizira da su L2. To su sledeći uzorci pod šiframa: MK5004 procjenjene starosti između 3.347-3.095 p.n.e.; MK5001, 3.348-3.035 p.n.e.; i SIJ002.A0101, starosti 3.349-3.033 p.n.e. Pored pomenutog rada L2 je nađen još u radu (Narasimhan, et al., 2018) gdje je identifikovan pod šifrom I2923, datirane starosti između 2.878-2.636 p.n.e., na lokalitetu Tepe Hisar u istočnom Iranu.
Kavkazka kultura Majkop prepoznata je kao kultura sa značajnim udjelom L2 haplogrupe, a kao što smo ranije naveli, njeni muški predstavnici pretežno su bili J2a-L26, J1, L1a, G2, L2, J2b temeljeći na rezultatima arheogenetike i današnje prisutnosti haplogrupa na području Kavkaza i okoline.
Najbliži zajednički predak L2 haplogrupe
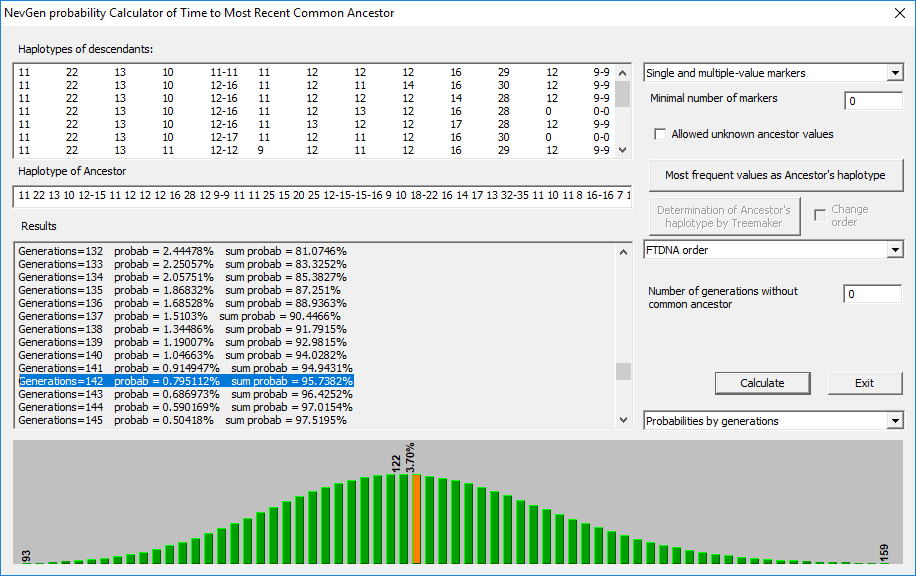
Za potrebe pisanja ovog rada identifikovali smo 15 haplotipova za koje se osnovano smatra ili je putem SNP testa potvrđena pripadnost L2 haplogrupi, a za koje ćemo kolektivnom kvadratnom metodom, metodom odstupanja od modala i uz pomoć NevGen Genealogy Tools v1.1 pokušati utvrditi vrijeme najbližeg zajedničkog pretka[1]. Za jednu generaciju prosječno smo uzeli 25 godina koliko se prema Devin (Devine, 2005) preporučuje pri računici koja broji više od 50 generacija do zajedničkog pretka. Pored vremena NZP-a utvrdićemo najsporije i najbrže markere za ovu haploskupinu. Kako bi smo stekli odgovarajuće okvire neophodne za dalju analizu.
Metodologija
Za proračun uzeto je 13 L2 haplotipova iz javne baze L haplogrupe-FTDNA (Y Haplogroup L Project, 2019) i to: N22419, 144196, N62831, N14510, 863058, 697332, 297364, 258180, 746307, 215799, 38049, 302190, N101576; te iz baze Srpskog DNK projekta, 103625 kao i jedan anonimni haplotip sa područja Zete. Korišćena je kolektivna kvadratna metoda; metoda odstupanja od modala; i računarski program NevGen Genealogy Tools v1.1. Za jednu generaciju uzet je prosjek od 25 godina. Srednja vrijednost STR za svih 15 haplotipova na Y67 markera je:
| DYS393 |
DYS390 |
DYS19 |
DYS391 |
DYS385 |
DYS426 |
DYS388 |
DYS439 |
| 11 |
22 |
13 |
10 |
12-15 |
11 |
12 |
12 |
| DYS389i |
DYS392 |
DYS389ii |
DYS458 |
DYS459 |
DYS455 |
DYS454 |
DYS447 |
| 12 |
16 |
28 |
12 |
9-9 |
11 |
11 |
25 |
| DYS437 |
DYS448 |
DYS449 |
DYS464 |
DYS460 |
YGATAH4 |
YCAII |
DYS456 |
| 15 |
20 |
25 |
12-15-16-17 |
9 |
10 |
18-22 |
16 |
| DYS607 |
DYS576 |
DYS570 |
CDY |
DYS442 |
DYS438 |
DYS531 |
DYS578 |
| 14 |
17 |
13 |
32-35 |
11 |
10 |
11 |
8 |
| DYF395S1 |
DYS590 |
DYS537 |
DYS641 |
DYS472 |
DYF406S1 |
DYS511 |
DYS425 |
| 16-16 |
7 |
11 |
10 |
8 |
12 |
9 |
12 |
| DYS413 |
DYS557 |
DYS594 |
DYS436 |
DYS490 |
DYS534 |
DYS450 |
DYS444 |
| 21-23 |
14 |
13 |
12 |
12 |
16 |
8 |
13 |
| DYS481 |
DYS520 |
DYS446 |
DYS617 |
DYS568 |
DYS487 |
DYS572 |
DYS640 |
| 25 |
22 |
14 |
12 |
11 |
14 |
10 |
11 |
Kvadratni metod, računica za NZP
Ovaj metod računice je najzahvalniji za sve vrste haplotipova i njihovu međusobnu kalkulaciju (Klyosov & Kilin, 2016). Kako mi u odabranoj skupini imamo haplotipove sa različitim brojem markera, ova metoda računice bila bi i najadekvatnija. Najbliži zajednički predak za svih 15 osoba prema KKK111 živio je prije 3.408±582 godina.
Odstupanje od modala, računica za NZP
Kako uzetih 15 haplotipova međusobno ima različit broj markera, te stoga imaju i različit broj međusobno uporedivih markera, za potrebe računice NZP metodom odstupanja od modala odredili smo standard Y17 (Yfiler) kao optilmalan; odnosno koji svi u većoj ili punoj mjeri ispunjavaju. Srednja vrijednost Y-STR na Y17 markera poređanih po FTDNA redosledu je:
| DYS393 |
DYS390 |
DYS19 |
DYS391 |
DYS385 |
DYS439 |
DYS389i |
DYS392 |
| 11 |
22 |
13 |
10 |
12-15 |
12 |
12 |
16 |
| DYS389ii |
DYS458 |
DYS437 |
DYS448 |
Y-GT4 |
DYS456 |
DYS438 |
DYS635 |
| 28 |
12 |
15 |
20 |
10 |
16 |
10 |
21 |
Uočena su 74 odstupanja od modala. Pri standardnoj stopi mutacije po haplotipu na standardu Y17 (Yfiler) od 0,034 dobijamo rezultat 145 generacija do zajedničkog pretka, odnosno ~3.627 godina. No kada izračunamo λ= prosječan broj mutacija po markeru korigovano za povratne mutacije i podjelimo sa konstantom brzine reakcije od 0,002; dobijamo rezultat od 169 generacija do zajedničkog pretka.
Odnosno ako uzmemo da je jedna generacija 25 godina dobijamo rezultat od ±4.225 godina do najbližeg zajedničkog pretka.
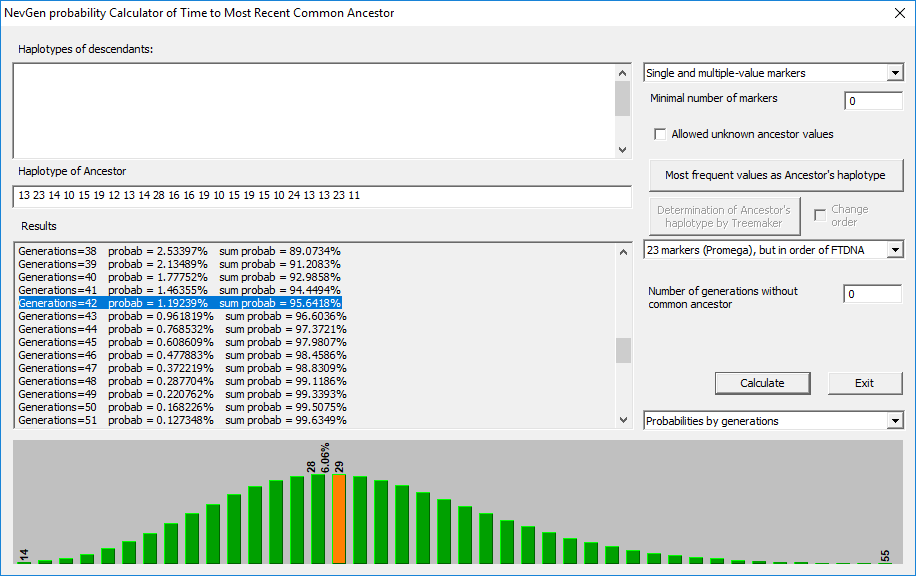
NevGen Genealogy Tools v1.1, računica za NZP
Prema ishodu računice na 95% vjerovatnoće, od zajedničkog pretka prošlo je 142 generacije, odnosno oko 3.550 godina.
![]()
Varijabilnost markera
Najveća varijabilnost na standardu Y17 bilježi se na markerima: DYS391, DYS385b, DYS439; DYS389ii; DYS456. Odnosno najstabilniji markeri su DYS393, DYS390, Y-GATA-H4, DYS438
Ishod
Dakle, proračun vremena NZP-a za svih 15 haplotipova kreće se od 3.408 do 4.225 godina. Odnosno predak L2 muškaraca živio je u periodu intezivnog uspona anadolskih kultura bronzanog doba.
Kako L2 izostaje u današnjoj populaciji Male Azije, Persije i Levanta, mala je vjerovatnoća da su učestvovali u populaciji Kura-Arakses kulture, pa bi migracione tokove L595 populacije smjestili u širi kontekst bronzanodopske Male Azije.
Antika i srednji vijek
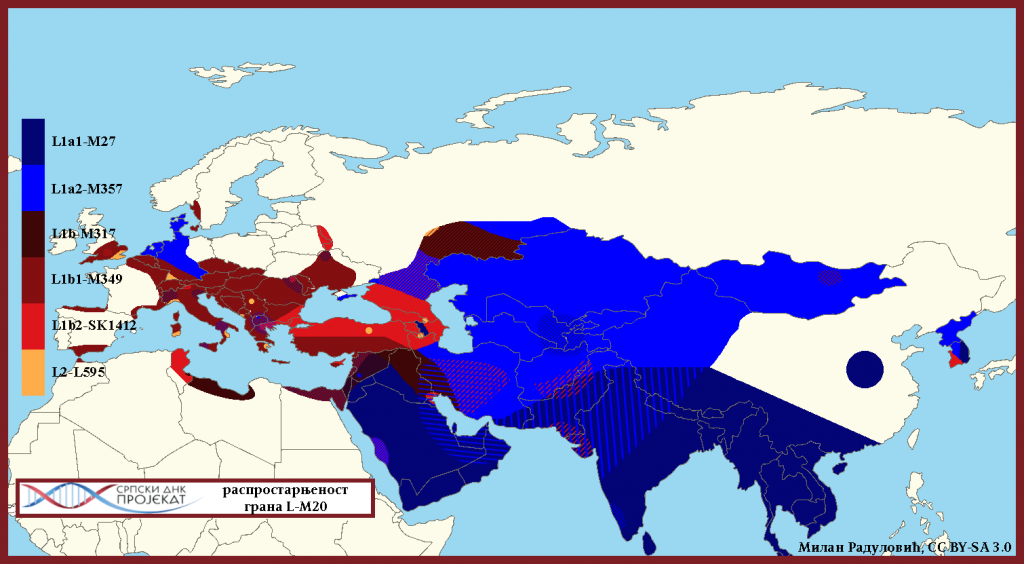
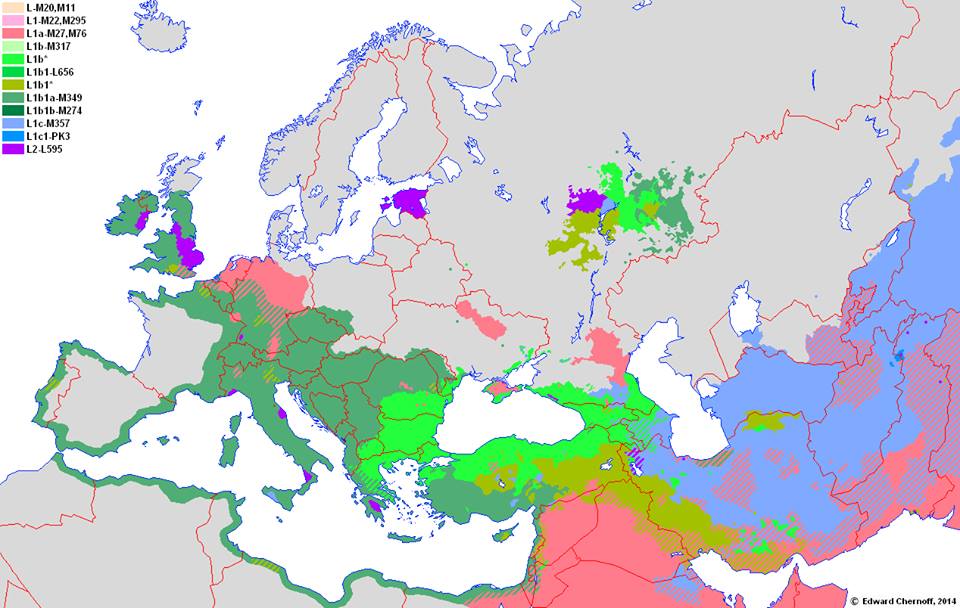
Gledajući u evropskim okvirima[2], haplogrupa L-M20 sreće se pretežno u granicama nekadašnjeg Rimskog carstva kao i J2-M172, ali s`tim da nema ravnomjerniju prisutnosti u granicama rimske države kao J2. Već je današnja koncentracija intezivna na nekadašnjoj granici Rimskog carstva, na koridoru rijeke Rajne.
U Evropi su mahom prisutni L1b haplotipovi, dok je u Danskoj i sjevernim dijelovima Njemačke prisutna i L1a, posebno M357 grana. U nedavno objavljenom radu o drevnim genomima iz vikinškog perioda (Margaryan, et al., 2019), dva od 300[3] analiziranih muških skeleta pripadali su L haplogrupi. Uzorak sa šifrom VK538 iz Apulije datiran u srednji vijek, pripadao je L1a-SK1426 haplogrupi, dok je uzorak pod šifrom VK355 iz Ševedske, Olanda, datiran u vikinško vrijeme[4] pripadao L2 haplogrupi.
Osim mediteranskih migracionih pravaca L haplogrupe prema i unutar Evrope, dio L-M20 mogao je stići i sa kretanjima avarskih i hunskih naroda, posebno imajući u vidu zastupljenost L1b-SK1412 u Dagestanu, te nalaz drevne DNK hunskog plemića iz Panonije, koji je pripadao L1a haplogrupi.
SNP mutacije
Haplogrupu L definišu snipovi M20, M11, M61, M185 i M295 (Karafet T. M., et al., 2008) ; ona se dijeli na tri grane L1a, L1b i L2:
- L1a definišu sledeće SNP mutacije: M2481, M2565, M2605, M2664, Y5525/Z20163, Y5560/Z20168
- L1b definišu sledeće SNP mutacije: M317/ FGC36877, L655, PH982, PH1071, PH1379, PH2312, PH2342, PH3435, PH3443, PH3615, PH3912, PH4035, PH4159, PH4188, SK1413
L2 definišu sledeće SNP mutacije: L595, L877/PF5780, Page6/PF5751
Tabela 1, osnovne grane i snip mutacije
| Y-DNK |
ISOGG |
SNP ekvivalenti |
| L-M20 |
L |
M20/PF5570 * M11 * M2437 * M2666 * M2607/S7544 * PF5790 * M2391 * Z20190/Y5564 * M2528 * PF5815/M2598 * PF5821/M2613 * PF5792/M2569 * PF5705/M2390 * PF5725/M2430 * PF5791 * PF5754/M2485 * PF5787/M2556 * PF5692/M2362 * M2552 * Y5530/Z20173 * M185/PF5755 * M2490 * PF5841/M2670 * M2464 * Y5521 * M2386 * M2381 * PF5757 * PF5563/M2597 * PF5756/M2492 * PF5829/M2640 * M2358 * M2530 * M2680 * M2418 * Y5522/Z20156 * M2650 * M2479 * M2401 * PF5717/M2407 * PF5726/M2435 * M2673 * PF5743/M2463 * PF5683/M2356 * M61/PAGE43/PAGES00043 * PF5729/M2444 * PF5532 * M2676 * Y5517/Z20124 * PF5810/M2587 * PF5813/M2595 * M2628 * M2594 * M2612 * PF5775/M2536 * PF5785/M2554 * M2669 * L878/PF5524 * PF5750/M2482 * M2457 * PF5695/M2370 * PF5693/M2368 * L855 * M2621 * L656 * M2436 * PF5773/M2532 * PF5723/M2426 * M2365 * Y5529/Z20172 * PF5715/M2404 * PF5528/M2403 * PF5737/M2453 * M2474 * PF5783/M2551 * PF5710/M2395 * M2642 * M2583 * M2366 * M2564 * PF5539 * Y5540/Z20112 * PF5830 * Y23631 * L863 * M2383 * |
| L1-M22 |
L1 |
M2494 * M295 * M2496 * M22 * M2614 * M2593 * M2546 * M2639 * Y5524/Z20161 * M2635 * Z20155 * M2600 * M2617 * M2517 * Y6264/Z20194 * M2471 * M2631 * PAGE121/PAGES00121 * CTS2285/M2465 * M2384 * M2419 * Y5520/Z20128 * M2445 * M2416 * Y5528/SK1410/Z20169 * Y5537/Z20202 * Y5534/Z20201 * Y5533/Z20200 * M2665 * M2420 * Y5545/Z20129 * M2460 * M2394 * M2399 * Z20160 * M2586 * CTS6368 * M2405 |
| L1a-M2481 |
L1a |
Y5525/Z20163 * M2605 * M2498 * M2375 * SK1418/Y5565 * M2565 * M2481 * Z20195/Y5566 * Z20185/Y5563 * M2560 * M2462 * M2545 * M2664 * Y5560/Z20168 * M2555 * M2568 * Z20182/Y5562 * M2480 * Y15954 * M2357 * Z20135 * M2431 * M2623 * M2520 * Y5544/SK1417 |
| L1a-M27 |
L1a1 |
Z20393 * Z20452 * Z8046 * Z20570 * Z20415 * M27 * Z20413 * Z20442 * Z20436 * Z20385 * Z20450 * Z20360 * Y15614 * Z20382 * Z8049 * Z20397 * Z20361/SK1421 * Z20437 * Z20365 * Z20362/SK1419 * Z20356 * Z20367 * Z20354 * Z20443 * Z20379 * Z20380 * Z20416/SK1422 * Z20363 * Z8050 * M76 * Z20449 * Z20418 * Z20392 * Z20359 * Z20419 * Z20433 * Z20435 * Z20364 * Z20358 * Z20369 * Z20383 * Z20429 * Z20425 * Z20384 * Z20336 * Z20405 * Z20438 * Z20406 * Z20445 * Z20371 * Z20381 * Z20424 * Z20368 * Z20402 * Z20394 * L1321 * Z8048 * Y15772 * Z20388 * Z20370 * Z20440 * Z20444 * Z20337 * Z20401 * Z20410 * Z20411 * Y20687 * Z20357 * Z20412 * Z20441 * Z20428 * L1319 * Z20451 * Z20446/SK1420 * Y17739 * Z8044 * Z20345 * Y20688 * Z20434 * Z20414 * Z20427 * Z20417 * Z20398 * Z20400 * Z20399 * Z20422 * Z20404 * Z24638/Z24638.2 * Z20407 * SK1423/SK1423.2 * Z20390/F11470 * Z20386 * P329/P329.1/P329.2 * Z20448 * S24646 * Y17738 * L430 |
| L1a-M357 |
L1a2 |
M2461 * Y5531/Z20252 * M2523 * M2659 * Y5535/FGC32242 * M2574 * Y5518/SK1446 * M2469 * Y5536/FGC32244 * Y5556/Z20249 * FGC32245/BY945 * M2542 * M2429 * M2632 * M2402 * M2360 * M2618 * M2627 * Y5548/Z20240 * M2514 * Y5551/Z20243 * M2592 * M2364 * L1307 * M2558 * M2540 * M2475 * M2549 * M2535 * M2533 * Z20232 * Y5527/Z20248 * M2432 * M2476 * M2451 * Y5558/Z20250 * M2610 * M2355 * Z20254/Y5532 * Y5515/Z20229 * M2380 * M2634 * Z20210/Y5539 * Z20237 * Y32086 * Z20241 * Z20263/Y12414 * M2507 * M2561 * M2679 * Z20221 * M2630 * M2575 * Z20228 * M2662 * M2581 * M2393 * Y125839 * M2412 * M2459 * Z20233 * Z20230 * M2439 * M2606 * Y5567/Z20256 * M2681 * M2441 * Z20223 * M2458 * M2653 * M2608 * Y5550/Z20242 * M2371 * Y5554/Z20247 * M2387 * Y5552/Z20244 * M2557 * M357 * Y5557/FGC32246 * M2601 * Y5526/Z20246 * Z20227 * M2359 * Y5559/SK1447/Z20251 * M2424 * FGC40731 * Z20238 * Z20226 * M2524 * M2529 * Z20236 * FGC32241/Y6263 * M2672 * Y6298/Z20231 * M2515 * M2361 * M2367 * M2671 * M2590 * Y20049* |
| L1b-M317 |
L1b |
Y16218 * Y16217/FGC36810 * Y16215/FGC36805 * Y16196/FGC36796 * PH3443 * PH3615 * SK1413 * Y16198/FGC36793 * Y16372/FGC36795 * Y16365/FGC36802 * PH4188 * Y16377/FGC36781 * Y16383/FGC36822 * S7560 * PH4035 * PH1379 * Y16376/FGC36828 * PH3435 * Y16202/FGC36804 * Y16197/FGC36811 * PH2312 * PH1071 * Y16195/FGC36783 * PH4159 * Y16361 * PH982 * M317 * Y16367/FGC36794 * FGC36800 * Y19618/FGC49074 * Y16201/FGC36797 * PH1736 * Y16388/FGC36816 * Y16393/FGC36775 * Y16213/FGC36825 * Y16391/FGC36786 * PH2342 * PH3912 * FGC36776 * Y18412 * Y19619/BY12538 * Y11005 |
| L1b-M349 |
L1b1 |
FGC36778 * BY12542 * FGC36830 * FGC36812 * BY12543 * FGC36779 * FGC36788 * FGC36820 * FGC36829 * FGC36826 * FGC36821 * FGC36818 * FGC36787 * FGC36785 * M349 * FGC36777 * FGC36815 * FGC36782 * FGC36813 * FGC36827 * FGC36791 * FGC36790 * FGC36806 * FGC36817 * FGC36801 * FGC42525 * FGC42542 * FGC42546 * FGC42556 * FGC42555 * FGC42551 * FGC36780 * FGC36809 * FGC36824 * FGC36808 * FGC36823 * FGC36789 * FGC36799 * FGC36814 * FGC36819 * FGC42531 * CTS8901/S6469 * FGC320 * PAGE113/PAGES00113 * Y2357/S24423/FGC7623 * FGC42535 |
| L1b-SK1412 |
L1b2 |
Y16390 * Y134788 * Y16370 * Y16395 * Y16385 * SK1412 |
| L2-L595 |
L2 |
L595 * L877 / PF5780 * PF5769 * PF5838 * PF5758 * PF5762 * L870 * PF5691 * PF5808 * PF5740 / Z20586 * PF5682 * PF5742 * Z20579 * Page6/PF5751 |
Nomenklatura
Tabela 2, sačinjena prema: (Rosser, et al., 2000; Scozzari, et al., 2001; The Y Chromosome Consortium, 2002; Karafet T. , et al., 2008; ISOGG, 2019).
| L-SNP |
Rosser et al. 2000 |
Scozzari et al. 2001 |
(α) |
(β) |
(γ) |
(δ) |
(ε) |
| L-M20 |
26 |
26.1/26.2 |
28 |
VIII |
1U |
27 |
Eu17 |
| L-M22 |
26 |
26.1/26.2 |
28 |
VIII |
1U |
27 |
Eu18 |
| L-M27 |
26 |
26.1/26.2 |
28 |
VIII |
1U |
27 |
Eu17 |
| L-M317 |
|
|
|
|
|
|
|
| L-M357 |
|
|
|
|
|
|
|
| L-L595 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| (ζ) |
(η) |
YCC 2002 (Longhand) |
YCC 2008 (Longhand) |
ISOGG 2006 |
ISOGG 2011 |
ISOGG 2015 |
|
| H5 |
F |
L* |
L |
L |
L |
L |
|
| H6 |
F |
L* |
L |
L |
L1 |
L1 |
|
| H5 |
F |
L1 |
L1 |
L1 |
L1a |
L1a1 |
|
|
|
|
L2 |
L2 |
L1b |
L1b |
|
|
|
|
L3 |
L3 |
L1c |
L1a2 |
|
|
|
|
|
|
L2 |
- |
|
skraćenice iz nomenklature:
α-Jobling and Tyler-Smith 2000 and Kaladjieva 2001; β-Underhill 2000;
γ-Hammer 2001; δ-Karafet 2001; ε-Semino 2000; ζ-Su 1999;
YCC-Y Chromosome Consortium;
ISOGG-International Society of Genetic Genealogy.
Mape rasprostranjenosti
![]()
![]()
![]()
Stablo
![]()
![]()
![]()
Podgrupe i zastupljenost
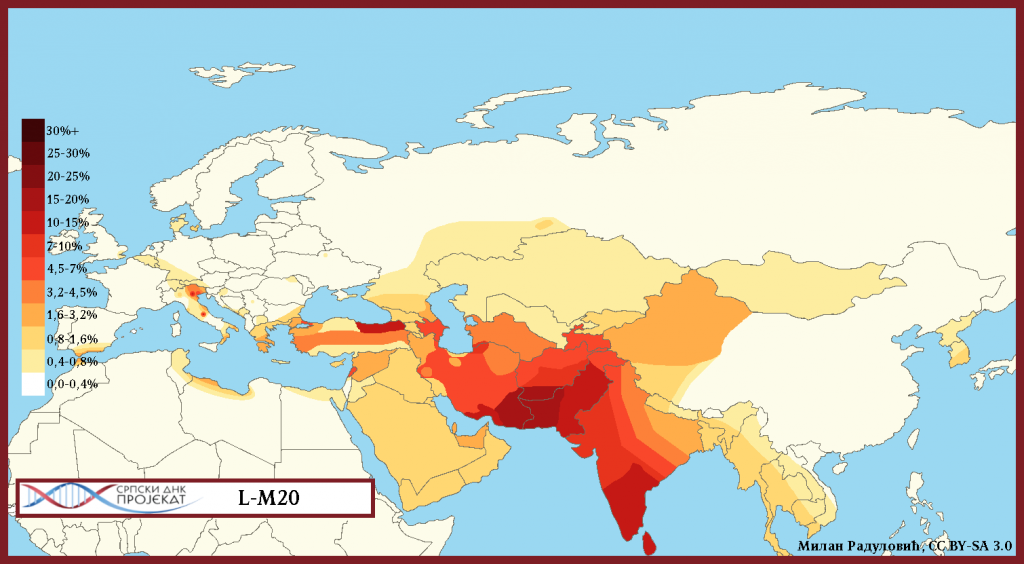
Haplogrupa L-M20 najzastupljenija je u južnoj Aziji, u Indiji, Pakistanu, Avganistanu. U Evropi je najveća koncentracija u sjevernoj Italiji, gdje je prema studiji iz 2009. (Battaglia, et al., 2009) zastupljena sa 4,5%. Procentualno gledano najzastupljenija je u zemljama: Šri Lanka 15%, Pakistan 13%, Avganistan 6,94%, Indija 6,73%, Iran 5%, Turska 4,2%.
| Procjena L-M20 populacije u navedenim zemljama |
| zemlja/narod/grupacija |
uzorak |
procenat |
±populacija |
| EVROPA |
933 |
0,1% (Dibirova, 2011) |
376.919 |
| Austrija-Tirol |
270 |
0,74% (Erhart, et al., 2012) |
2.761 |
| Belgija-Antverpen, Istočna Flandrija |
540 |
0,74% (2012; Decorte, et al., 2012) |
6.456 |
| Bugarska |
808 |
0,2% (Karachanak, et al., 2013) |
6.845 |
| Grčka |
257 |
2,33% (Greek DNA Project , 2019) |
121.394 |
| Estonija |
74 |
1,35% (Scozzari, et al., 2001) |
8.384 |
| Italija |
817 |
0,24% (Grugni, et al., 2018) |
70.626 |
| Kipar |
344 |
1,2% (Heraclides, et al., 2017) |
5.058 |
| Mađarska |
535 |
0,37% (Hungarian_Magyar_Y-DNA_Project, 2019) |
17.284 |
| S. Makedonici |
428 |
0,46% (Noveski, Trivodalieva, Efremov, & D., 2010; Jankova, Geppert, Videtic Paska, Willuweit, & Roewer, 2019; Macedonian DNA, 2019; Srpski DNK projekat, 2019) (Battaglia, et al., 2009) |
4.600 |
| Srbi |
2.997 |
0,13% (Srpski DNK projekat, 2019) |
7.085 |
| Holandija |
85 |
1,17% (Karafet, et al., 2016) |
99.766 |
| Hrvatska |
1.100 |
0,27% (Mršić, et al., 2012) |
5.351 |
| Španija-Andaluzija |
29 |
3,4% (Semino, et al., 2000) |
142.598 |
| EVROPA UKUPNO |
|
0,13%, minimalna procjena |
498.208 |
| AZIJA |
|
|
|
| Avganistan |
433 |
6,94%[5] (Di Cristofaro, et al., 2013) |
1.094.873 |
| Azerbejdžan |
72 |
6,9% |
325.024 |
| Gruzija |
66 |
1,5% (Battaglia, et al., 2009) |
32.280 |
| Indija |
728 |
6,73% (Sengupta, et al., 2006) |
42.343.511 |
| Irak |
203 |
0,99% (Abu-Amero, et al., 2009) |
166.830 |
| Iran |
|
5% (Grugni, et al., 2012) |
1.890.300 |
| Jermenija |
413 |
1,93% (Herrera, et al., 2012) |
30.002 |
| Katar |
72 |
2,78% (Abu-Amero, et al., 2009) |
26.952 |
| Koreja |
506 |
0,59% (Kim, Han, Kim, & Kim, 2010) |
215.769 |
| Kina-Sinkjang |
546 |
2% (Hou, 2019) |
218.133 |
| Kirgistan |
132 |
1,5% (Di Cristofaro, et al., 2013) |
40.860 |
| Liban |
916 |
5,24% (Abu-Amero, et al., 2009) |
112.450 |
| Malezija-Indijci |
301 |
17,27% (Chang, et al., 2008) |
173.788 |
| Mongolija |
933[6] |
0,5% (Dibirova, 2011) |
7.606 |
| Pakistan |
176 |
13% (Sengupta, et al., 2006) |
13.505.349 |
| Palestina |
367 |
0,7% (Zalloua, et al., 2008) |
15.926 |
| Saudijska Arabija |
157 |
1,91% (Abu-Amero, et al., 2009) |
315.150 |
| Sirija |
202 |
5,4% (Zalloua, et al., 2008) |
460.751 |
| Tadžikistan |
142[7] |
6,33% |
282.666 |
| Turska |
523 |
4,2% (Cinnioğlu, et al., 2004) |
1.722.081 |
| Uzbekistan |
170 |
0,56% (Völgyi, et al., 2014) |
93.111 |
| UAE |
164 |
3,06% (Abu-Amero, et al., 2009) |
140.846 |
| Šri Lanka |
60 |
15% (Karafet, et al., 2016) |
1.625.250 |
| AFRIKA |
|
|
|
| Egipat |
117 |
0,85% (Karafet, et al., 2016) |
412.424 |
| Libija |
173 |
1,73% (Immel, 2008) |
62.280 |
| Tunis |
220 |
0,45% (Fadhlaoui-Zid, et al., 2014) |
24.710 |
| UKUPNO |
|
|
65.837.130 |
Sledeće grane gravitiraju:
L-M20>L595>
- Vrlo rijetka haplogrupa. Za sada je nađena kod svega nekoliko desetina testiranih, i to u: Sardiniji, Srbiji, Grčkoj, Italiji, Turskoj, Njemačkoj, Rusiji, Engleskoj, Irskoj i Azerbejdžanu.
L-M20>M22>M2481>M27>
- Ova grana L1a haplogrupe najprisutnija je u južnoj Aziji, Indiji i Pakistanu; a prepoznatljiva je i u Arabijskom poluostrvu. Bilježi se i u Kini i na Levantu; a ima i svoje evropske grane koje se nalaze pretežno u mediteranskom pojasu. Grana se na dvije grane, Z20387 i Y31961 (dominantna grana Y31214).
- Z20387 karakteristična je za arabijsko poluostrvo i južnu Aziju. Dalje se grana na dvije velike grane: BY12397 koju za sada nalazimo na Arabijskom poluostrvu i L1320 koja je pretežno južnoazijskog karaktera, no koja ima i svoje arabijske i levantske grane.
- Y31961 dijeli se na Y31961* i Y31213. Grana Y31213 zastupljena je od Levanta preko Arabije do centralne Azije.
L-M20>M22>M2481>M2533>M357>
- Prepoznatljiva je centralnoazijska grana L1a haplogrupe. Ova grana ima jako stare grane i u Arabiji i Levantu. Grana se na četiri grane: M2398 (M2398>Y12415 gravitira Tadžikistanu), Y6288, Y44848 (Arabijsko poluostrvo) i BY198125.
L-M20>M22>M2481>M2533>M357>Y6288>Y6259>Y6284>Y6266
- Y6266 je grana koja je karakteristična za Čečene, kod kojih je L1a-M357 zastupljena sa 7,2%. Starost grane je ±3.300 godina.
L-M20>M22>M317>
- L1b je grana koja ima svoje matično područje na prostoru Zakavkazja i sjevera Iraka, odakle se tokom bronzanog doba proširila u Evropu. Do zajedničkog pretka broji oko 12.500 godina. Grana se na dvije grane SK1412 i M349.
L-M20>M22>M317> SK1412
- Gravitira Pontu i Anadoliji, što je i bazna teritorija haplorupe, zajedno sa Zakavkazjem; prostor na kome se haploskupina i razvila. Prisutna je od Evrope i Mediterana do centralne i istočne Azije. Grana se na dvije velike grane FGC51074 i Y16366.
- FGC51074 raširena je od istočnog Mediterana do Britanije. Sa Levantom kao matičnim podrujem.
- Y16366 je pontsko-kavkaska grana, koje se dijeli na Y16187 i Y16366*.
L-M20>M22>M317> SK1412> Y16366
- Pontsko-kavkaska grana dijeli se na Y16187 i Y16366*:
- Y16187 je jedna od prepoznatljivih haplogrupa Grka sa Ponta i iz Anadolije, gdje je L1b-M317 prisutna sa oko 17% od ukupne muške populacije. Nizvodna Y18413>Y18891 pored Ponta prisutna je u Gruziji i među Avarima.
- Y16366* identifikovana je na širokom području između istočne Anadolije i Kavkaza.
L-M20>M22>M317>M349
- Raširena je na prostoru Evrope iduž Mediterana. Sreće se i u kavkaskom regionu. Za sada su identifikovane tri nizvodne grane: B374, FGC36845 i Y31183.
- B374 nazvan još i Rajna-Dunav-Volga klaster, po matičnom području haploskupine, klaster je iksljučivo evropski.
- FGC36845 raširen je diljem Evrope, Mediterana, a sreće se i na Levantu.
- Y31183 mahom se sreće na Levantu, ali je takođe kao i svi ogranci M349 prisutna na području Evrope.
- Kod nas je vodeća haplogrupa od L-M20, čije se matično područje na prostoru srpske zemlje nalazi na potezu Trebinje-Bjelopavlići-Grbalj.
Stari narodi
L-M20 može se povezati sa starim narodima Anadolije i njihovim državama, Hetitima i Lidijcima; sa levantskim narodima Feničanima i Filestejcima; Persijancima; Dravidima i drugim sa područja južne Azije; ali i sa Hunima i Avarima.
Filestejci
Zaključujući prema rezultatima studije „Ancient DNA sheds light on the genetic origins of early Iron Age Philistines“ i nalzu ASH087.A0101[8] koji je L haplogrupe, utvrđeno je da su učestvovali u etnogenezi Filestejaca (Feldman, et al., 2019). Prema rezultatu iz pomenutog rada, utvrđeno je da pomenuti Filestejac i njegov klaster ASH_IA2 nemaju evropske primjese za razliku od grupe ASH_IA1 kod kojih je ona oko 14%, što ukazuje na lokalno levantsko porijeklo i ono stanovništvo koje se inkorporiralo u filestejsku populaciju[9].
Feničani
Sasvim je izvjesno da su učestvovali u etnogenezi Feničana na šta upućuje prethodno pomenuti rad koji svjedoči o prisustvu populacije na Levantu, ali i pojačano prisustvo u regijama u kojima su Feničani bili aktivni, kao što su Andaluzija, sjever Afrike, jug Italije.
Hetiti i drugi anadolski narodi
Prema današnjoj i nedavnoj (prije 1922.g.) demografskoj slici; odnosno rasprostranjenosti, učestalosti (4-12%) i raznovrsnosti L1 haploskupine na području istočne i sjeveroistočne Anadolije; sasvim je izvjesno da su i L-M20 muškarci imali udjela u drevnim civilizacijama Anadolije. Posebno ukazavši na brojne arheogenetske nalaze sa područja kavkaskog regiona iz koga su tekli pravci migracija ka Anadoliji.
Persijanci
Nesumnjivo je potvrđena autohtonost haploskupine na području Irana gdje su arheogenetski rezultati dodatno potvrdili starost skupine. U Iranu su ravnomjerno raspoređeni na cijeloj teritoriji što ukazuje na rano učešće u etnogenetskim procesima.
Huni
Kako je u svom radu Dibirova navela, zastupljenost haplogrupe na prostoru Evroazijske stepe je 0,5% (Dibirova, 2011) što je malo, ali je arheogenetskim nalazom iz Mađarske br. 12763 (Džansugurova, i dr., 2017) halogrupa L potvrđena kod Huna. Nađena je u pomenutom uzorku koji je pripadao hunskom plemiću. Uzorak datiran u ±500. godinu. Rezultatom je utvrđena zastupljenost haplogrupe kod Huna, ali je ostala zagonetka u kom omjeru je učestvovala u hunskoj populaciji. Iz uzorka su izvučeni sledeći STR markeri, razvrstali smo ih po FTDNA redosledu:
| DYS393 |
DYS19 |
DYS385 |
DYS439 |
DYS389i |
DYS389ii |
| 16 |
14 |
7-8 |
19 |
10 |
24 |
| DYS458 |
DYS437 |
YGATAH4 |
DYS456 |
DYS438 |
DYS635 |
| 14 |
16 |
7 |
15 |
12 |
20 |
Na osnovu haplotipa možemo zaključiti da se radi o L-M20>M22. Bližu granu sa većom sigurnošću ne možemo identifikovati, ali se na osnovu markera uočava okvirna sličnost sa M357 haplotipovima. Prije svega niske vrijednosti DYS385 markera koje se sreću isključivo kod M357. Većina predviđača Y-DNK haplogrupe ne može prepoznati haplogrupu na osnovu haplotipa, dok „World Haplogroup & Haplo-'I' Subclade Predictor“, za haplovrstu predviđa L3 odnosno M357.
Srpske zemlje
U srpskim zemljama identifikovane su sledeće haplogrupe: L1b-M349>BY12542, L2-L595 koje su mahom stacionirane u Crnoj Gori/Primorju; i L1a-M357 koja gravitira Povardarju.
L1b-M349>BY12542, Dobrogorani
Najbrojniji je klaster L haploskupine u srpskoj zemlji, često ga još nazivamo i Lazarevići, po bratstvu iz čijeg su se okruženja razivla sva druga bratstva. Matična oblast roda je Primorje. Prema analizi Vukićevića, stručnjaka za genetičku genealogiju Crne Gore, razvoj i migracije BY12542 bratstava išli su (Vukićević, 2019):
- Lazarevići su bratstvo iz Grblja koje potiče iz Šobajića u Bjelopavlićima. Od njih su Vujići i Pejurići u Bjelopavlićima;
- Od iseljenika iz Šobajića su bratstva: Vučurovići, Vodovari i Ilići u Krivošijama i Vukalovići i Vučinići u Dolovima u Grahovu koja su migrirala preko Zagarača i Dobre Gore u Cucama, a po kojoj su prozvani Dobrogoranima.
- U Zagaraču su ostali Radmanovići, razgranata grupa bratstava, koji se rođakaju sa Dobrogoranima, a ova veza je i genetski potvrđena.
- Dio iseljenika iz Šobajića zaustavio se na Krivošijama i od njih potiču Vukalovići i Vujičići u Zubcima i Grahovu i Miloševići u oblasti Gacka i Nevesinja.
Prema Vukićeviću (Vukićević, 2019B): „na osnovu predanja o srodstvu i poreklu pojedinih bratstava iz Šobajića u Bjelopavlićima osnovano se može pretpostaviti da je prisutna i u Bjelopavlićima, Grblju, Ljubotinju, Piperima, Ceklinu i Cucama.“
Jedna od karakteristika roda je i krsna slava Sv. Petka koju slavi većina roda, njih oko 80%. U inostranim i naučnim bazama podataka nemaju ni bliža ni daleka poklapanja. Administratori „L - Y Haplogroup L“ (Y Haplogroup L Project, 2019) svrstavali su srpski klaster u veći libanski klaster.
Najbliži zajednički predak
Za potrebe pisanja ovog rada identifikovali smo haplotipove iz baze Srpskog DNK projekta na dan 29. juli 2019. godine, za koje se osnovano smatra ili je putem SNP testa potvrđena pripadnost BY12542 haplogrupi. Za koje ćemo kolektivnom kvadratnom metodom, metodom odstupanja od modala i uz pomoć NevGen Genealogy Tools v1.1 pokušati utvrditi vrijeme najbližeg zajedničkog pretka. Generaciju prosječno računamo 30 godina.
Metdologija
Korišćena je kolektivna kvadratna metoda; metoda odstupanja od modala; i računarski program NevGen Genealogy Tools v1.1. Za jednu generaciju uzet je prosjek od 30 godina. Srednja vrijednost STR za sve identifikovane haplotipova na Y23 markera je:
| DYS393 |
DYS390 |
DYS19 |
DYS391 |
DYS385a |
DYS385b |
DYS439 |
DYS389i |
| 13 |
23 |
14 |
10 |
15 |
19 |
12 |
13 |
| DYS392 |
DYS389ii |
DYS458 |
DYS437 |
DYS448 |
Y-GH4 |
DYS456 |
DYS576 |
| 14 |
28 |
16 |
16 |
19 |
10 |
15 |
19 |
| DYS570 |
DYS438 |
DYS481 |
DYS549 |
DYS533 |
DYS635 |
DYS643 |
| 15 |
10 |
24 |
13 |
13 |
23 |
11 |
Kvadratni metod, računica za NZP
Ovaj metod kako smo ranije naveli je najzahvalniji za sve vrste haplotipova i njihovu međusobnu računicu (Klyosov & Kilin, 2016). Kako mi u odabranoj skupini imamo haplotipove sa različitim brojem markera, ova metoda računice bila bi i najadekvatnija. Najbliži zajednički predak za sve osobe prema KKK111 živio je prije 427±158 godina.
Odstupanje od modala, računica za NZP
Uzeti haplotipovi ispunjavaju Yfiler standard, stoga smo za potrebe računice NZP metodom odstupanja od modala odredili Y17 (Yfiler) kao optilmalan, odnosno koji svi u punoj mjeri ispunjavaju.
Uočena su 22 odstupanja od modala. Pri standardnoj stopi mutacije po haplotipu na pri standardu Y17 (Yfiler) od 0,034 dobijamo rezultat 40 generacija do zajedničkog pretka, odnosno ~1.200 godina. Kada izračunamo λ= prosječan broj mutacija po markeru korigovano za povratne mutacije i podjelimo sa konstantom brzine reakcije od 0,002; dobijamo rezultat od 41,5 generacija do zajedničkog pretka.
Odnosno ako uzmemo da je jedna generacija 30 godina dobijamo rezultat od ±1.245 godina do najbližeg zajedničkog pretka.
NevGen Genealogy Tools v1.1, računica za NZP
Prema ishodu računice na 95% vjerovatnoće od zajedničkog pretka prošlo je 42 generacije, odnosno oko 1.200 godina.
![]()
Ishod
Proračun vremena NZP-a za sve uzete haplotipova kreće se od 430 do 1.200 godina zavisno od računača. Dvije od tri računice poklapaju se u ishodu, odnosno prema njima najbliži zajednički predak živo je prije 1.200 godina. Dakle negdje u 9. vijeku. Usled malog broja markera i relativne malojbojnosti grupe, treba biti obazriv pri uzimanju rezultata.
Razmatranje
Dobrogorani se mogu posmatrati kroz kontekst bratstava porijeklom iz Šobajića u Bjelopavlićima, a koja imaju bliske, naizgled matične veze sa bratstvom Lazarevića iz Grblja.
U pogledu praporijekla za sada ne možemo dati konkretne odgovore. Prije svega jer nemamo identifikovanu/testiranu nizvodnu granu od BY12542 iz koje proističe rod, nadalje nemamo reprezentativan broj testiranih sa većim brojem markera što bi nam omogućilo kvalitetniju NZP računicu. Rod nema poklapanja sa drugim L1b haplotipovima i nema bliskih klastera unazad hiljadama godina, što dodatno otežava rekonstrukciju migracije ka Balkanu. No, kako je bio svrstan u libanski klaster od strane administratora L haplogrupe na FTDNA[10], to nam daje indiciju o istočnom porijeklu grupe, a zajedno sa starošću od 1.200 godina smiješta ga u grko-romejski supstrat srpskog etnosa.
Stara DNK
Do sada je najstariji pronađeni uzorak L haploskupine datiran u eneolit 4.350-3.500 p.n.e., I1407, nađen u Areni pećine na prostora Jermenije. Arheogenetski nalazi mahom su nađeni na prostoru kavkaskog regiona i Baktrijsko-Margianskog arheološkog kompleksa. U tabeli koja slijedi, navedeni su svi dosadašnji nalazi L haploskupine:
Tabela 3, drevna L
| šifra |
datovanje |
prostor |
država |
Y-DNK |
mt-DNK |
studija |
| I1634 |
4.330-4.060 p.n.e. |
Areni pećina |
Jermenija |
L1a1-M27 |
H2a1 |
(Lazaridis, et al., 2016) |
| I1632 |
4.230-4.000 p.n.e. |
Areni pećina |
Jermenija |
L1a1-M27 |
K1a8 |
- |
| I1407 |
4.350-3.500 p.n.e. |
Areni pećina |
Jermenija |
L1a1-M27 |
H |
- |
| DA85 |
169-250 n.e. |
Kurgan br. 2, Žaparik |
Kirgizija |
L1a1-M27>Y31961* |
|
(Damgaard, et al., 2018) |
| I5604 |
1880-1697 p.n.e. |
Bustan |
Uzbekistan |
L1a |
K1a1 |
(Narasimhan, et al., 2018) |
| I4285 |
1873-1661 p.n.e |
Sapali Tepe |
Uzbekistan |
L1a |
U7a3 |
- |
| S10001.E1.L1 |
1300-1000 p.n.e. |
Loe Banr 1 |
Pakistan |
L1a |
R30b1 |
- |
| I8220 |
970-550 p.n.e. |
Aligrama |
Pakistan |
L1a |
.. |
- |
| I6555 |
906-820 p.n.e. |
Loe Banr 1 |
Pakistan |
L1a |
M4 |
- |
| S10974.Y1.E1.L1 |
900-800 p.n.e. |
Loe Banr 1 |
Pakistan |
L1a |
.. |
- |
| I6554 |
831-796 p.n.e. |
Loe Banr 1 |
Pakistan |
L1a |
T2g1 |
- |
| S7721.E1.L1 |
500-300 p.n.e. |
Svat dolina, Sajdu Šarif |
Pakistan |
L1a |
R6b |
- |
| I2923 |
2878-2636 p.n.e. |
Tepe Hisar |
Iran |
L2 |
W3b |
- |
| MK5004 |
3347-3095 p.n.e. |
Marinskaja 5 |
Rusija |
L2 |
T2al |
(Wang, et al., 2018) |
| MK5001 |
3348-3035 p.n.e. |
Marinskaja 5 |
Rusija |
L2 |
K1a4 |
- |
| SIJ002.A0101 |
3349-3033 p.n.e. |
Sinjuha |
Rusija |
L2 |
U4c1 |
- |
| VK355 |
847 ± 65 n.e. |
Oland |
Švedska |
L2 |
U5b1b1a |
(Margaryan, et al., 2019) |
| VK538 |
11-13. vijek |
Fođa |
Italija |
L1a1b-SK1426 |
H+16291 |
- |
| 12763 |
500 n.e. |
Panonska nizija |
Mađarska |
L1a2-M357 |
D4j12 |
(Džansugurova, i dr., 2017) |
Poznate ličnosti
Seid Adžal Šemsudin
Seid Adžal Šemsudin ( 1211 , Buhara - 1279 ) prvi je guverner carske dinastije Juan u provinciji Junan. Učesnik je mongolskih osvajanja u Burmi, a poznat je po tome što je predak mnogim muslimanima širom Kine. Haplogrupa L1a1-M27 (Wang, et al., 2013)
![]() Dženg He
Dženg He
Čeng He (trad. kin. 鄭和, kin. 郑和; 1371—1435) bio je kineski admiral, pomorac i istraživač. Istraživao je područja jugoistočne azije, Sumatre, Jave, Šri Lanke, Indije, Persije, Persijskog zaliva, jugozapadne Azije, Crvenog mora, i Mozambika. Nadimak mu je San Bao, pa se spekuliše o tome da se po njemu nazvao Sindbad. Potomak je čuvenog Adžal Šemsudina. Haplogrupa L1a1-M27.
![]() Luka Vukalović
Luka Vukalović
Luka Vukalović (Zupci, Trebinje, 18. oktobar 1823 — Saltakča, Odesa, Rusija 6. jul 1873) je vođa srpskog ustanka u Hercegovini 1852—1862. Knjaz Danilo imenovao je Luku za vojvodu Zubaca, Kruševice i Dračevice. Haplogrupa L1b-M349>BY12542.
![]() Tripko Vukalović
Tripko Vukalović
Tripko Vukalović (Zubci, Trebinje, 1835 -Nikšić, 1. april 1898) je jedan od ustaničkih vođa tokom Nevesinjske puške, vojvoda je od Zubaca i najmlađi brat vojvode Luke Vukalović. Haplogrupa L1b-M349>BY12542 rod Dobrogorana.
Literatura:
Vukićević, I. (2019, jun 20). Rodovi starobalkanskog porekla u Crnoj Gori: dopunjeno 20.6.2019. Preuzeto sa Poreklo: https://www.poreklo.rs/2017/11/19/rodovi-starobalkanskog-porekla-u-crnoj-gori/
Vukićević, I. (2019B, jul 14). Genetska slika Crne Gore: dopunjeno 14.7.2019. Preuzeto sa Poreklo: https://www.poreklo.rs/2017/11/19/genetska-slika-crne-gore/
Džansugurova, L. B., Džantaeva, K. B., Nuržibek, Žunusova, G. S., Kuzovleva, E. B., Musralina, L. Z., . . . Husainova, Э. M. (2017). Vыdelenie i analiz drevneй DNK iz kostnыh čelovečeskih ostankov gunnskogo perioda. Seriя biologičeskaя i medicinskaя, 4(322), 39-50.
Dibirova, H. D. (2011). Rolь geografičeskoй podrazdelennosti i lingvističeskogo rodstva v formirovanii genetičeskogo raznoobraziя naseleniя Kavkaza (po dannыm ob Y hromosome). Moskva: Učreždenii Rossiйskoй akademii medicinskih nauk Mediko-genetičeskom naučnom centre RAMN .
Radulović, M., & Novaković, N. (2018). Haplogrupa J2 (sa osvrtom na srpske zemlje). Beograd: Društvo srpskih rodoslovaca Poreklo.
Rajevac, M. (2019). Haplogrupa L-drevna DNK. Beograd: Društvo srpskih rodoslovaca "Poreklo": Srpski DNK projekat.
Srpski DNK projekat. (2019, Jul 23). Srpski DNK projekat. Preuzeto sa Poreklo: https://dnk.poreklo.rs/DNK-projekat
Abu-Amero, K. K., Hellani, A., González, A. M., Larruga, J. M., Cabrera, V. M., & Underhill, P. A. (2009). Saudi Arabian Y-Chromosome diversity and its relationship with nearby regions. BMC Genetics, 1-9. doi:10.1186/1471-2156-10-59
Alizadeh, K., Eghbal, H., & Samei, S. (2015). Approaches to social complexity in Kura-Araxes culture: A view from Köhne Shahar (Ravaz) in Chaldran, Iranian Azerbaijan. Paléorient, 41(1), 37-54.
Battaglia, V., Fornarino, S., Al-Zahery, N., Olivieri, A., Pala, M., Myres, N. M., & Semino, O. (2009). Y-chromosomal evidence of the cultural diffusion of agriculture in southeast Europe. European Journal of Human Genetics, 820-830. doi:10.1038/ejhg.2008.249
Chang, Y. M., Jaya, P., Revathi, P., Phoon, Y., Jaya, P., & Jeevan, N. (2008, July 3). Population Malaysia [Indian]. Retrieved from YHRD: https://yhrd.org/YP000242-Malaysia+%5BIndian%5D
Cinnioğlu, C., King, R., Kivisild, T., Kalfoglu, E., Atasoy, S., Cavalleri, G., . . . Underhill, P. (2004). Excavating Y-chromosome haplotype strata in Anatolia. Human Genetics, 127-148. doi:10.1007/s00439-003-1031-4
Damgaard, P. d., Marchi, N., Rasmussen, S., Peyrot, M., Renaud, G., Korneliussen, T., . . . Pedersen, M. W. (2018). 137 ancient human genomes from across the Eurasian steppes. Nature, 557(7705), 369–374 . doi: 10.1038/s41586-018-0094-2
Decorte, R., & al., e. (2012, February 10). Population Oost-Vlaanderen, Belgium [Belgian]. Retrieved from YHRD: https://yhrd.org/YP000784-Oost-Vlaanderen%2C+Belgium+%5BBelgian%5D
Decorte, R., Larmuseau, M., & Ottoni, C. (2012, February 10). Population Antwerpen, Belgium [Dutch]. Retrieved from YHRD: https://yhrd.org/YP000781-Antwerpen%2C+Belgium+%5BDutch%5D
Devine, D. (2005, September/October). How Long Is a Generation? Ancestry Magazine, 23(5), 51-55. Retrieved from Ancestry: https://books.google.ba/books?id=JzgEAAAAMBAJ&lpg=PA1&hl=sr&pg=PA51#v=onepage&q&f=false
Di Cristofaro, J., Pennarun, E., Mazières, S., Myres, N. M., Lin, A. A., Temori, S. A., & Chiaroni, J. (2013). Afghan Hindu Kush: where Eurasian sub-continent gene flows converge. PloS one, 8(10), 1-12. doi:10.1371/journal.pone.0076748
Erhart, D., Berger, B., Niederstätter, H., Gassner, C., Schennach, H., & Parson, W. (2012, August 17 ). Frequency data for 17 Y-chromosomal STRs and 19 Y-chromosomal SNPs in the Tyrolean district of Reutte, Austria. International Journal of Legal Medicine, 126(6), 977–978. doi:doi.org/10.1007/s00414-012-0751-1
Fadhlaoui-Zid, K., Garcia-Bertrand, R., Alfonso-Sánchez, M. A., Zemni, R., Benammar-Elgaaied, A., & Herrera, R. J. (2014). Sousse: extreme genetic heterogeneity in North Africa. Journal of Human Genetics, 1-9. doi:10.1038/jhg.2014.99
Feldman, M., Master, D. M., Bianco, R. A., Burri, M., Stockhammer, P. W., Mittnik, A., . . . Krause, J. (2019). Ancient DNA sheds light on the genetic origins of early Iron Age Philistines. Science Advances, 5(7), 1-10. doi:10.1126/sciadv.aax0061
Greek DNA Project . (2019, July 21). Greek DNA Project - Y-DNA Classic Chart. Retrieved from Family Tree DNA: https://www.familytreedna.com/public/Greece?iframe=yresults
Grugni, V., Battaglia, V., Hooshiar Kashani, B., Parolo, S., Al-Zahery, N., & al., e. (2012, July 18). Ancient Migratory Events in the Middle East: New Clues from the Y-Chromosome Variation of Modern Iranians. PLOS ONE. doi:10.1371/journal.pone.0041252
Grugni, V., Raveane, A., Mattioli, F., Battaglia, V., Sala, C., Toniolo, D., . . . Semino, O. (2018). Reconstructing the genetic history of Italians: new insights from a male (Y-chromosome) perspective. Annals of human biology, 44-56. doi:10.1080/03014460.2017.1409801
Hay, M. (2018, September 28). Haplogroup J2. Retrieved from Eupedia: https://www.eupedia.com/europe/Haplogroup_J2_Y-DNA.shtml
Heraclides, A., Bashiardes, E., Fernández-Domínguez, E., Bertoncini, S., Chimonas, M., & Christofi, V. (2017). Y-chromosomal analysis of Greek Cypriots reveals a primarily common pre-Ottoman paternal ancestry with Turkish Cypriots. PLOS ONE, 12(6). doi:doi.org/10.1371/journal.pone.0179474
Herrera, K. J., Lowery, R. K., Hadden, L., Calderon, S., Yepiskoposyan, L., Regueiro, M., . . . Herrera, R. J. (2012). Neolithic patrilineal signals indicate that the Armenian plateau was repopulated by agriculturalists. European Journal Of Human Genetics, 20, 313-320. doi:10.1038/ejhg.2011.192
Hou, Y. (2019). Y-SNP Branch Information on L. Retrieved from YHRD: https://yhrd.org/tools/branch/L
Hungarian_Magyar_Y-DNA_Project. (2019, July 29). Family Tree DNA - Hungarian_Magyar_Y-DNA_Project. Retrieved from Family Tree DNA: https://www.familytreedna.com/public/Hungarian_Magyar_Y-DNA_Project?iframe=yresults
Immel, U. D. (2008, July 3). Population Tripoli, Libya [Libyan]. Retrieved from YHRD: https://yhrd.org/YP000416-Tripoli%2C+Libya+%5BLibyan%5D
ISOGG. (2019, July 31). Y-DNA Haplogroup Tree. Retrieved from ISOGG (International Society of Genetic Genealogy): https://isogg.org/tree/
Jankova, R., Geppert, M., Videtic Paska, A., Willuweit, S., & Roewer, L. (2019). Y-chromosome diversity of the three major ethno-linguistic groups in the Republic of North Macedonia. Forensic Science International: Genetics. doi:doi.org/10.1016/j.fsigen.2019.07.007
Karachanak, S., Grugni, V., Fornarino, S., Nesheva, D., Al-Zahery, N., Battaglia, V., . . . Semino, O. (2013). Y-Chromosome Diversity in Modern Bulgarians: New Clues about Their Ancestry. PLoS ONE, 8(3), 1-9. doi:10.1371/journal.pone.0056779
Karafet, T. M., Bulayeva, K. B., Nichols, J., Bulayev, O. A., Gurgenova, F., Omarova, J., . . . Hammer, M. F. (2016). Coevolution of genes and languages and high levels of population structure among the highland populations of Daghestan. Journal of Human Genetics, 181–191. doi:10.1038/jhg.2015.132
Karafet, T. M., Mendez, F. L., Meilerman, M. B., Underhill, P. A., Zegura, S. L., & Hammer, M. F. (2008). New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree. Genome research, 18(5), 830-838. doi:10.1101/gr.7172008
Karafet, T., Mendez, F., Meilerman, M., Underhill, P., Zegura, S., & Hammer, M. (2008). New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree. Genome Research, 830-839. doi:10.1101/gr.7172008
Kim, S. H., Han, M. S., Kim, W., & Kim, W. (2010, November). Y chromosome homogeneity in the Korean population. International Journal of Legal Medicine, 124(6), 653–657. doi:doi.org/10.1007/s00414-010-0501-1
Klyosov, A. A., & Kilin, V. V. (2016). Kilin-Klyosov TMRCA Calculator for Time Spans up to Millions of Years. Advances in Anthropology, 6(3), 51-71. doi:http://dx.doi.org/10.4236/aa.2016.63007
Kohl, P. L. (2007). The Making of Bronze Age Eurasia. New York: Cambridge University Press.
Lazaridis, I., Nadel, D., Rollefson, G., Merrett, D. C., Rohland, N., Mallick, S., . . . Reich, D. (2016). Genomic insights into the origin of farming in the ancient Near East. Nature, 2016/07/25/online. doi:10.1038/nature19310
Macedonian DNA. (2019, July 23). Macedonian DNA. Retrieved from Family Tree DNA: https://www.familytreedna.com/groups/macedonia/about/background
Mahal, D. G., & Matsoukas, I. G. (2017). Y-STR Haplogroup Diversity in the Jat Population Reveals Several Different Ancient Origins. Frontiers in Genetics, 8:121. doi:10.3389/fgene.2017.00121
Margaryan, A., Lawson, D., Sikora, M., Racimo, F., Rasmussen, S., Moltke, I., . . . de Barros Damgaard, P. (2019). Population genomics of the Viking world. bioRxiv. doi:10.1101/703405
Mršić, G., Petek, M., Vrdoljak, A., Popović, M., Valpotić, I., Anđelinović, Š., . . . Primorac, D. (2012). Croatian national reference Y-STR haplotype database. Molecular Biology Reports, 39, 7727–7741. doi: 10.1007/s11033-012-1610-3
Narasimhan, V. M., Patterson, N., Moorjani, P., Lazaridis, I., Lipson, M., Mallick, S., . . . Coppa, A. (2018). The Genomic Formation of South and Central Asia. bioRxiv. doi:10.1101/292581
Noveski, P., Trivodalieva, S., Efremov, G., & D., P.-K. (2010). Y chromosome single nucleotide polymorphisms typing by SNaPshot MINISEQUENCING. Balkan Journal of Medical Genetics, 13(1), 1-13. doi:10.2478/v10034-010-0013-9
Rosser, Z., Zerjal, T., Hurles, M., Adojaan, M., Alavantic, D., & Amorim, A. (2000). Y-Chromosomal Diversity in Europe Is Clinal and Influenced Primarily by Geography, Rather than by Language. Am J Hum Genet, 67(6), 1526-43.
Sahoo, S., Singh, A., Himabindu, G., Banerjee, J., Sitalaximi, T., Gaikwad, S., . . . Kashyap, V. K. (2006, January 24). A prehistory of Indian Y chromosomes: Evaluating demic diffusion scenarios. PNAS, 103(4), 843–848. doi:10.1073/pnas.0507714103
Scozzari, R., Cruciani, F., Pangrazio, A., Santolamazza, P., Vona, G., & Moral, P. (2001). Human Y-chromosome variation in the western mediterranean area: Implications for the peopling of the region. Human Immunology, 871-884. doi:doi.org/10.1016/S0198-8859(01)00286-5
Semino, O., Passarino, G., Oefner, P. J., Lin, A. A., Arbuzova, S., Beckman, L. E., . . . Francalacci, P. (2000). The Genetic Legacy of Paleolithic Homo sapiens sapiens in Extant Europeans: A Y Chromosome Perspective. Science, 290(5494), 1155-1159. doi:10.1126/science.290.5494.1155
Sengupta, S., Zhivotovsky, L. A., King, R., Mehdi, S. Q., Edmonds, C. A., Chow, C. E., & Underhill, P. A. (2006). Polarity and Temporality of High-Resolution Y-Chromosome Distributions in India Identify Both Indigenous and Exogenous Expansions and Reveal Minor Genetic Influence of Central Asian Pastoralists. The American Journal of Human Genetics, 78, 202–221. doi:10.1086/499411
The Y Chromosome Consortium. (2002). A Nomenclature System for the Tree of Human Y-Chromosomal Binary Haplogroups. Genome Research, 12(2), 339–348. doi:10.1101/gr.217602
Trivedi, R., Sahoo, S., Singh, A., Bindu, G. H., & Banerjee, J. (2007). High Resolution Phylogeographic Map of Y-Chromosomes Reveal the Genetic Signatures of Pleistocene Origin of Indian Populations. Anthropology Today: Trends, Scope and Applications, 393-414.
Völgyi, A., Zalán, A., Szvetnik, E., Nagy, M., & Pamzsav, H. (2014, October 31). Population Uzbekistan. Retrieved from YHRD: yhrd.org
Wang, C.-C., Reinhold, S. R., Kalmykov, A., Wissgott, A., Brandt, G., & al., e. (2018, May 16). The genetic prehistory of the Greater Caucasus. BioRxiv. doi:10.1101/322347
Wang, C.-C., Wang, L.-X., Zhang, M., Yao, D., Jin, L., & Li, H. (2013). Present Y chromosomes support the Persian ancestry of Sayyid Ajjal Shams al-Din Omar and Eminent Navigator Zheng He. arXiv, 1-5. Retrieved from https://arxiv.org/ftp/arxiv/papers/1310/1310.5466.pdf
Y Haplogroup L Project. (2019, June). L - Y Haplogroup L. Retrieved from Family Tree DNA: https://www.familytreedna.com/groups/y-haplogroup-l/about
YFull. (2019, April 2). YFull YTree v7.02.01. Retrieved from YFull: https://www.yfull.com/tree/L/
Zalloua, P. A., Xue, Y., Khalife, J., Makhoul, N., Debiane, L., & Platt, D. E. (2008). Y-Chromosomal Diversity in Lebanon Is Structured by Recent Historical Events. Am J Hum Genet, 873–882. doi:10.1016/j.ajhg.2008.01.020
[1] Najbliži zajednički predak, u daljem tekstu i skraćeno kao NZP.
[2] Posmatrajući Evropu u gegrafskom smislu.
[3] Ukupno je analizirano 442 skeleta, od čega je 300 muških.
[4] 847 ± 65 nove ere.
[5] Etnički sastav Avganistana i unutar etnički odnosi L-M20 haplogrupe su sledeći: 42.1% Paštuni, 7,5%→518.516; 33.6% Tadžici, 6,33%→349.271; 10.6% Uzbeci, 9,44%→164.322; 9.8% Hazari, 3,9%→ 62.764; 3.9% Ostali →Nepoznato.
[6] Prema rezultatu Dibirove na 933 uzorka evroazijske stepe, a koja uključuje i Mongoliju, zastupljenost L-M20 haplogrupe je 0,5%.
[7] Prema rezultatima Tadžika iz Avganistana, zastupljenost L-M20 haplogrupe je 6,33%.
[8] Jedan od 3 rezultata iz kojih je izvučen Y hromozom.
[9] Filestejci su po pisanju narod za koji se sugeriše kulturna veza s mikenskim svijetom u Grčkoj i indoevropski jezik.
[10] Prema stanju iz februara 2019. godine.
The post Haplogrupa L-M20: sa osvrtom na srpske zemlje appeared first on Poreklo.




































 Dženg He
Dženg He Luka Vukalović
Luka Vukalović Tripko Vukalović
Tripko Vukalović